By T. W. Sherry and Cody M. Kent
Linked Paper: Extensions and limitations of MacArthur (1958): A review of ecological and evolutionary approaches to competition and diet in the New World wood warblers (Parulidae) by T.W. Sherry and C.M. Kent. Ornithology.
Explaining the variety of avian species requires understanding a variety of species interactions, in both ecological and evolutionary time scales. In particular, competition among species is a prominent, if battle-scarred, interaction. Our recent review paper employs several new approaches and information, especially dietary data, to address warbler competition and coexistence, which we examine from both ecological and evolutionary perspectives. Our study contributes to a Special Feature: Advances in Avian Diet Methods and Applications in the American Ornithological Society journal, Ornithology.

Ecologists got the jump on explaining why there are so many species with ecological coexistence mechanisms focused on concepts including the ecological niche, interspecific competition, and resource partitioning. MacArthur’s (1958) Ph.D. dissertation was a seminal contribution using spruce woods warblers, and follow-up research, that helped launch decades of studies on resource partitioning and coexistence—and controversy about the prevalence of competition in nature. However, despite this body of research, the central question still remains: Do coexisting competitor species generally “partition” resources? And, if not, then how does interspecific competition work?
We argue in our paper that interspecific competition is indeed pervasive in warbler communities, necessary to our understanding of species coexistence. However, an important, and largely overlooked, mechanism is diffuse exploitation involving many species competing simultaneously and depressing resource abundance. This mechanism has selected evolutionarily, we argue, for diversification and specialization not in the resources—including insects and spiders—on which the largely insectivorous warblers feed and overlap substantially (and thus fail to partition), but rather in the substrates where they feed. Substrates are not resources, are not used up by consumers, and thus are not “partitioned.” Instead, the slight phenotypic differences of coexisting species, including their beaks, wings, tarsi, and tails, appear to make a difference in how efficiently these birds use different substrates harboring difficult-to-exploit prey, helping the predators survive through periods of food scarcity.

MacArthur’s foundational 1958 paper has greatly influenced subsequent thought on coexistence, including his documented foraging differences of coexisting warblers, differences that likely accessed slightly different prey. However, he was handicapped by not knowing what his study species ate—still largely unknown for breeding spruce-woods warblers! Other information unavailable to MacArthur, and that emphasizes why we need to continue to integrate ecological and evolutionary perspectives, is avian phylogenies. For example, Lovette and Hochachka (2006) suggest why coexisting warblers are not each other’s closest relatives phylogenetically; and the Oliveros et al. (2019) passerine phylogeny helps explain warblers’ origins and insinuation into Neotropical communities. More recent work, including André Dhondt’s book, Interspecific Competition in Birds (Oxford University Press, 2012), provided us a critical insight. Dhondt identified the kinds of evidence needed to demonstrate interspecific competition, much of which, we realized, we already possessed based on long-term demographic studies of long-distance migrant parulid warblers wintering in Jamaica. What we lacked was detailed diet data, controlling for location and timing, since insect abundances typically vary spatially and temporally.
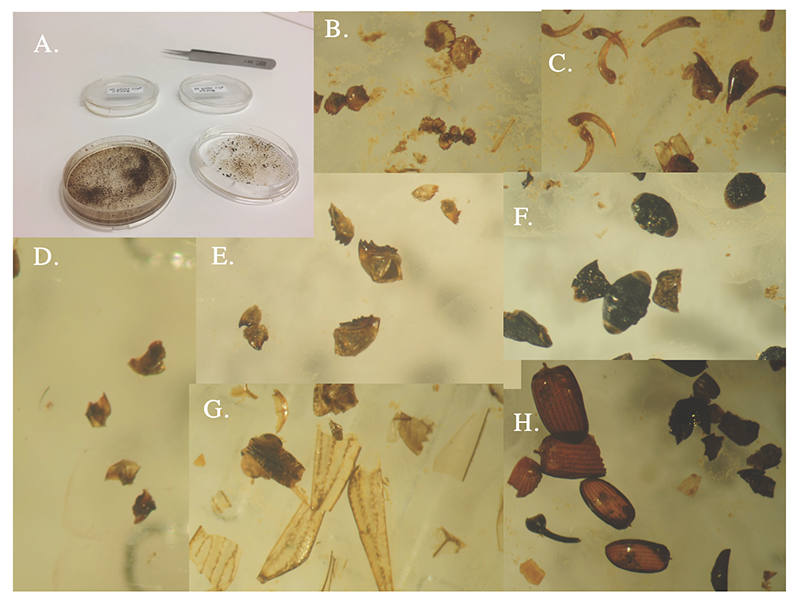
Once we obtained such dietary data from a variety of Jamaican (and breeding) habitats, we could make a strong inferential case for diffuse exploitative competition, without the necessity of field experiments, which are challenging considering warbler mobility and local diversity. Another insight came once we were lab-bound, mucking through Jamaican warbler stomach contents and foraging substrate data. We realized that evolutionarily equivalent bird species occupy mainland Neotropical habitats, largely representing taxa that never made it to the Caribbean Islands, including most suboscine passerines. This dearth of competitors suggests an ecological opportunity on Caribbean Islands that helped fuel the striking warbler adaptive radiation. Moreover, poor island-colonizing ability by many mainland insectivorous birds may have resulted from a feeding specialization-dispersal tradeoff (Sherry et al. 2020). After reviewing and synthesizing these kinds of evidence, we find that warbler species interactions are far more complex than generally appreciated. Much work remains to rethink how competition over different time scales shapes communities in this family of birds that has been so foundational to our understanding of species interactions.
Wellv done! Always interesting to see how influential those classic papers stand the test of time.
Alan, Thanks! I really enjoyed writing this paper, both honoring MacArthur’s enormous contributions to the field, but also looking forward to potentially novel and productive ways to look at the same question(s) MacArthur addressed!